Introduction
Low-intensity resistance training combined with blood-flow restriction (LIBFR) induces significant benefits in terms of muscle strength, explosiveness and hypertrophy (Gepfert et al., 2020; Wilk et al., 2018, 2020a). While there is some evidence that LIBFR increases muscle size to a similar extent as high-intensity (HI) resistance training (Loenneke et al., 2012), it generally induces less gain in muscle strength compared to HI (Hughes et al., 2017; Lixandrao et al., 2018; Loenneke et al., 2012). Nevertheless, despite being performed at much lower intensity, the magnitude of the acute responses is typically greater for LIBFR than for HI. For instance, a number of studies using different exercise protocols reported a greater decline in muscle strength after acute LIBFR rather than HI exercise (up to 37% vs. up to 19%, respectively) (Cook et al., 2007, 2013). It has also been shown that LIBFR leads to pronounced impairment in contractile function (Husmann et al., 2018). However, not all studies observed such magnitude of decline in force production immediately after LIBFR. As it has been recently shown, when acute exercise is performed with BFR and set for a predefined number of repetitions, the decrement in maximal voluntary contraction (MVC) is similar between LIBFR and HI conditions (Fatela et al., 2017; Kim et al., 2017). Ultimately, this may well explain the inconsistent findings of past research on the interaction between both training modalities and muscular fatigue. In addition, it also indicates that variations in the prescription of LIBFR can be used to fine-tune the development of fatigue (quantified via decrements in MVC). Although this measure is considered as the most valid and reliable procedure to assess muscle fatigue, its functional value has been recently questioned (Maffiuletti et al., 2016; Rodriguez-Rosell et al., 2018). In fact, in many sports and daily activities the available time to develop force is typically lower than that necessary to produce maximal force (Aagaard et al., 2002).
The rate of torque development (RTD), which represents the ability to increase torque as quickly as possible during a rapid voluntary contraction (Δ torque/Δ time), is important for motor actions performed explosively. This is particularly relevant in circumstances when torque production times are short (e.g. 100-300ms), such as when reversing a fall or in various athletic events (e.g. sprints, throws and jumps) (Aagaard et al., 2002). Nonetheless, the effect of fatigue on the ability to produce torque rapidly has received less attention. Some studies have shown that the impingement of fatigue on the RTD occurs differently than on MVC, indicating that the acute changes in muscle strength and the RTD are not necessarily associated (Buckthorpe et al., 2014). Accordingly, fatigue exerts a more rapid and profound effect on explosive torque production than on MVC, particularly during the initial 50ms of contraction. This is associated with impairment in the ability of the central nervous system to maximally activate the muscle during the first milliseconds (0-50ms) of contraction (Morel et al., 2015). From then on (>50ms), reductions in the RTD are more strongly related with changes in intrinsic contractile properties and lower muscle strength (Folland et al., 2014). Exploring acute neuromuscular fatigue induced by different modalities of resistance exercise would help understand the observed adaptation subsequent to specific training programs. Thus, it is important to gain a more comprehensive insight into the differential effect of both these hypertrophic-resistance training approaches on muscle fatigue. Ultimately, the combined analysis of both the absolute and relative RTD (normalized to maximal strength) might unravel the degree to which the impact of HI exercise and LIBFR on performance may share common etiological factors (i.e. reduction in muscle strength or changes in other mechanisms).
We aimed at investigating the influence of HI exercise and LIBFR on the development of explosive torque throughout the torque-time curve. The sequential RTD was measured to discriminate the eventual role of each exercise protocol on specific determinants of explosive torque production. It was hypothesized that RTD would respond similarly to HI exercise and LIBFR.
Methods
Participants
Seventeen healthy, young and moderately active Sport and Exercise Science students (8 men and 9 women, age: 22.0±1.9 yrs, body height: 167.5±8.9 cm, body mass: 63.8±9.5 kg; BMI 22.7±1.9 kg/m2) were included. All participants were active to a similar extent, accumulating 9 h of physical activity per week as part of their academic work. Moreover, they were well accustomed to resistance training and strength testing procedures (as part of their classes). All participants had a normal body mass index, were normotensive (Whelton et al., 2018) and free from known diseases. This study was approved by the Faculty’s Ethics Committee (CEFMH Nº 4/2017) and complied with the Declaration of Helsinki. At study entry, participants were also required to complete a health-screening questionnaire and a degree of handedness questionnaire (Elias et al., 1998) and provide written informed consent to participate. Participants were all tested with the dominant upper-limb.
Design and Procedures
Participants attended three laboratory sessions scheduled for the morning period between 09:00 and 11:00 am. Participants did not engage in intense physical exercise for 24 h before testing. The first session was designed to measure the elbow-flexion one-repetition maximum (1RM) and to familiarize all participants with BFR exercise. Anthropometric and blood pressure measurements were also taken for each participant. Finally, all participants practiced several isometric MVCs (System 3, Biodex Medical Systems, Shirley, NY) and submaximal elbow flexion with and without BFR (at 20 and 75% 1 RM, respectively) at the end of the first visit. Afterwards, and using a randomized, counterbalanced within-subjects design, participants underwent two experimental conditions (HI exercise vs. LIBFR), separated by a 1-wk period. On the remaining two laboratory visits, each participant performed isometric MVCs before and immediately after exercise (HI resistance exercise vs. LIBFR). This enabled the subsequent quantification of changes in peak torque and the RTD resulting from acute resistance training. Exercise was initiated 15 min after baseline MVC measurements and involved dumbbell elbow flexion performed in the seated position for both protocols. Resting arterial occlusion pressure (AOP) was determined immediately before LIBFR. This was done to set exercise blood flow restriction at 60% of AOP.
Measurements
One-repetition maximum testing
1RM was determined for elbow flexion of the dominant upper limb using dumbbells (the same free weights used during acute resistance exercise), while the participant remained seated on an adjustable chair. The back of the arm was firmly planted against the pad of the chair during all lifting attempts. Testing procedures followed the recommendations advanced by the American Society of Exercise Physiologists (Brown and Weir, 2001). Briefly, the protocol was initiated with 8-10 repetitions performed at a load of ~ 50% of estimated 1RM. Then, the load was adjusted to 75% of the estimated 1RM and each participant was instructed to complete five repetitions. These two initial sets of muscle contractions served as a specific warm up. Subsequently, the load was adjusted as necessary and each participant performed the maximum number of repetitions until volitional failure (with verbal encouragement). The load was increased by 5% whenever participants were able to complete more than two repetitions before failure. Three minutes of rest were allowed between the first three trials. Further trials required 5 min of rest to ensure full recovery (Harris et al., 1976). A repetition was considered valid if the participant lifted the dumbbell through the entire range of motion (from full elbow extension to maximum elbow flexion). Before attempting each concentric lift, participants were asked to pause for 1 s at the end of the eccentric phase. 1RM was accepted as the maximum load that each participant could mobilize in single maximum dynamic elbow flexion. All participants received strong verbal encouragement during each attempt and 1RM was always determined within 5 attempts.
Resting arterial occlusion pressure
As performed in past research, all blood-flow measurements were taken during seated rest (Wilk et al., 2020b). The arterial blood flow was detected using a vascular Doppler probe (SONOLINE B LCD Fetal Doppler 8MHz vascular probe, CONTEC, China), placed over the radial artery, at the wrist level. Pulse was detected via auditory and visual signals obtained from the Doppler probe. A narrow 6 x 83cm pneumatic nylon cuff (SC5 Tourniquet Cuffs, D. E. Hokanson, Inc. Bellevue, WA) was placed on the most proximal end of the dominant arm and inflated using a rapid inflation device (E20 Rapid Cuff Inflator, D.E. Hokanson, Inc. Bellevue, WA). Inflation was initially set to 50% of the individual resting systolic blood pressure and then raised gradually up to the point when radial pulse was interrupted. AOP was recorded as the nearest 1 mmHg pressure at which pulse was not present.
Elbow flexion maximal voluntary isometric contractions
After performing 10-12 repetitions through full range of motion using the dynamometer isokinetic mode and being securely fixed with chest and waist straps, participants’ MVC was tested in the seated position at an angle corresponding to 600 of elbow flexion using the isometric mode. Participants were instructed to avoid any countermovement prior to MVC and to exert their maximum force as fast and hard as possible throughout the full duration of each trial (Holtermann et al., 2007). Verbal encouragement was provided to each participant. MVC was recorded as peak torque (N⋅m) obtained in three isometric contraction trials lasting 3 s each. One min of recovery was allowed between trials. Past research indicates that this protocol is reliable for testing the RTD at different time epochs (Holtermann et al., 2007). The highest torque value observed at this specific time point was defined as pre-exercise MVC. For both conditions, post-exercise MVCs were always taken within 30 s of the final repetition of the last set of acute resistance training. No discernible countermovement or pre-tension (change in baseline torque during the 100 ms prior to contraction onset) were identifiable in the trials used for analysis.
Resistance exercise
The exercise load was set at 75 and 20% 1RM for HI and LIBFR sessions, respectively. LIBFR was performed at 60% of resting AOP. HI resistance exercise involved 4 sets of 10 repetitions (2 s concentric; 2 s eccentric), with 60s of rest between sets (Ratamess et al., 2009). LIBFR was prescribed for 4 sets of 30+15+15+15 repetitions (1 s concentric; 1 s eccentric) with 30 s recovery between sets. This protocol was prescribed for elbow-flexion exercise and has been used in studies examining the physiological responses to BFR resistance training (Loenneke et al., 2013). BFR was achieved by inflating the nylon cuff within the most proximal portion of the arm. Spotting was provided to the participants whenever they were unable to successfully lift the weight. Finally, cuff pressure was released after LIBFR immediately before the post-exercise isometric MVC.
Rate of torque development analysis
The dynamometer torque signal was sampled at a 1000Hz analog-to-digital conversion rate by an external analog-to-digital converter (National Instruments, USB-6251). Then, the signal was smoothed offline using a digital fourth-order, zero-lag Butterworth filter with a cutoff frequency of 10 Hz. Additionally, before testing, torque was corrected for the effect of gravity on the upper limb in accordance with the manufacturer recommendations.
Two different investigators, following previous recommendations, visually identified signal onsets of each contraction (Tillin et al., 2010, 2013). The RTD (Δ torque/Δ time) was quantified as the average slope of the torque-time curve for 3 sequential 50-ms time periods (RTD0-50, RTD50-100 and RTD100-150). According to past research, a sequential RTD assessed over discrete consecutive time periods isolates any changing contributions of neural and contractile determinants throughout the time course of contraction (Folland et al., 2014). Torque and sequential RTDs were expressed in absolute terms and relative to MVC obtained at each time point. Determining the relative RTD is important, because the absolute RTD is influenced by peak force capacity of the muscles (Andersen et al., 2005). Conversely, a normalized RTD enables comparisons of the ability to utilize the available MVC torque in different conditions (e.g. before and after a fatiguing exercise task) (Maffiuletti et al., 2016). Finally, we also computed the peak RTD as the steepest part of the curve using a moving sampling window of 20 ms (Haff et al., 2015).
Statistical Analysis
Before comparing both conditions (LIBFR vs. HI resistance exercise), data were tested for normality and homoscedasticity with Kolmogorov-Smirnov and Mauchly’s tests, respectively. Based on a pilot study including 6 participants (3 men and 3 women), when the true difference between both conditions (HI exercise vs. LIBFR) in decreasing the peak RTD corresponded to 130.8±123.9 N⋅m⋅s-1, a sample size of 17 participants was estimated to have more than 80% power of correctly rejecting the null hypothesis.
Since the levels of fatigue resulting from different regimens of acute resistance exercise vary as a function of the volume load (i.e. nº sets x nº repetitions x load lifted) (Loenneke et al., 2011), we explored whether the volume load was similar between LIBFR and HI protocols (using paired t-tests). The effect of each exercise protocol on changing each dependent variable was determined using a two-way repeated measures ANOVA with two within-subject factors [(condition: LIBFR vs. HI resistance exercise) x (time: pre- vs. post-exercise)]. The eta-squared values (proportion of total variance that is attributable to an effect) are reported for significant findings (small, medium and large effect: 0.01, 0.06 and 0.14, respectively) (Cohen, 1988). The Hedges’s g was also reported to indicate effect sizes (ES) (small, medium and large effect: 0.2, 0.5 and 0.8, respectively) (Cohen, 1988). The intraclass correlation coefficients (ICC) were additionally calculated for pre-exercise data with the purpose of exploring the between-sessions reliability for MVC and the peak RTD. The ICC values varied between 0.71 and 0.88 which represents good reliability (Cicchetti and Sparrow, 1981). All statistical analyses were computed using the Statistical Software Package for Social Sciences (SPSS 24.0, SPSS Inc., Chicago, IL) and significance was set at p<0.05. All data are reported as means ± SD.
Results
Participants’ 1RM was equivalent to 14.0±4.7 kg. AOP taken before LIBFR was equivalent to 136.5±13.8 mmHg. The volume load was higher for HI exercise (427.1±134.9 kg) than LIBFR (213.6±67.3 kg) (p<0.0001). MVC peak torque was similar between conditions at baseline (pre-HI: 42.4±17.3 vs. pre-LIBFR: 41.2±12.7 N⋅m; ICC: 0.88, p<0.0001). There were no differences between protocols in eliciting a significant decrement in MVC peak torque after exercise (HI: -28.0±17.6 vs. LIBFR: -21.7±13.3%) (time main effect: F=37.2, p<0.0001, η2:0.69, ES:-0.39; condition x time interaction: F=1.8, p=0.69). As with peak torque, we found that both conditions (i.e. HI exercise and LIBFR) caused a significant reduction in the peak RTD from pre- to post-exercise (pre-HI exercise: 792.6±578.8, post-HI exercise: 428.8±218.1; pre-LIBFR: 802.1±459.6, post-LIBFR: 430.6±234.5 N⋅m⋅s-1) (time main effect: F=19.1, p<0.0001, η2:0.55, ES:-0.81 and -0.99; respectively). Importantly, the magnitude of this effect was similar between HI and LIBFR conditions (condition x time interaction: F=0.04; p=0.83). The ICC between pre-exercise values for the peak RTD corresponded to 0.71 (p=0.001).
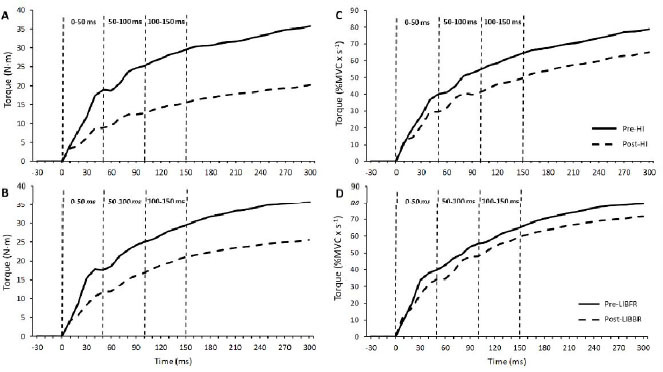
As can be seen in Figure 1, the steepest portion of the torque-time trace was attained during the first 50 ms of the curve. Then, there was a progressive decline in the RTD over time and this occurred similarly between conditions (condition x time interaction, p>0.05). Significant main effects of time were obtained for the absolute RTD quantified between 0-50 (F=16.9, p=0.001, η2:0.51, ES:1.99) and 50-100ms (F=25.0, p<0.0001, η2:0.61, ES:2.45) (Table 1).
Table 1
Changes in absolute and normalized sequential rate of torque development after high-intensity (HI) and low-intensity blood-flow restricted (LIBFR) resistance exercise.
[i] Values are mean ± SD and 95% confidence intervals. MVC, isometric maximal voluntary contraction. Comparisons between protocols were made with repeated measures analysis of variance with two within-subject factors [(condition: LIBFR vs. HI resistance exercise) x (time: pre- vs. post-exercise)]. * Significant differences between pre- and post-exercise time points (p < 0.05)
Conversely, after normalizing torque values to MVC obtained at each time point, this was only sustained for RTD0-50 (time main effect: F=6.5, p=0.02, η2:0.29, ES:1.24) (Table 1). Under these circumstances, RTD50-100 remained essentially unchanged from pre- to post-exercise in both conditions (F=3.5, p=0.13).
Figure 1
Mean value of torque production recorded in all participants during maximal isometric contraction of the elbow flexor muscles before and after acute high-intensity (HI) exercise and low-intensity resistance training combined with blood-flow restriction (LIBFR). Absolute torque production at (A) pre- and (B) post-exercise time points. Relative torque production at (C) pre- and (D) post-exercise time points. The torque signal was analyzed in three successive 50-ms time windows from their respective onsets (0-50, 50-100 and 100-15 ms).
Discussion
To our knowledge, this is the first study to determine the effects of acute LIBFR on the RTD. Moreover, we also explored whether both LIBFR and HI resistance exercise induced a similar decline in the RTD at different time epochs. In agreement with that hypothesized, we found that both conditions elicited similar effects on the sequential RTD computed during the first 150ms of MVC (0-50, 50-100 and 100-150ms). Specifically, while the absolute RTD decreased consistently up to 100ms post-exercise, the relative RTD remained essentially unchanged beyond the first 50ms of rising muscle torque following contraction onset. Thus, our findings indicate that the decline in elbow flexion MVC post-HI exercise and LIBFR is greatly responsible for the attenuation of the RTD between 50-100ms of isometric contraction. In contrast, this is not the case for the first 50ms of torque production. Finally, it should be emphasized that these results were obtained in a context of different volume loads between protocols (the volume load of HI exercise almost doubled that of LIBFR).
Despite being considered independent variables that express different functional and neuromuscular abilities (de Ruiter et al., 2008), several studies have reported parallel changes in the RTD and isometric MVC with acute muscle fatigue (Molina and Denadai, 2012; Penailillo et al., 2015). In addition, a positive association exists between the RTD and maximal muscle strength, especially for the RTD obtained in the latter phase of rising torque (i.e., beyond 75 ms of contraction) (Andersen and Aagaard, 2006; Folland et al., 2014; Maffiuletti et al., 2016). Yet, such a relationship between MVC and the absolute RTD has been reported as relatively weak (Holtermann et al., 2007). Thus, it is believed that the absolute RTD and relative RTD reflect different neuromuscular qualities. Ultimately, the relative RTD provides a unique opportunity to discriminate between different physiological mechanisms influencing the expression of the RTD independently of the reductions in MVC resulting from acute muscle fatigue (Waugh et al., 2014). For this reason, we explored the impact of each mode of resistance exercise on the sequential values of the relative RTD and compared it with that obtained for the absolute RTD.
We found that both exercise regimens induced a pronounced rightward shift of the torque-time curve. This is supported by a significant reduction in peak and sequential absolute RTD values after exercise. The analyses of the sequential RTD revealed that, while the rightward displacement of absolute torque production was extensive to the first 100ms of contraction, the effects of fatigue on the decreasing relative RTD were only seen during the initial 50ms of rising muscle torque. Therefore, our data indicate that the decrement in MVC resulting from LIBFR and HI exercise appears to be largely responsible for lower explosive torque production (i.e. decreased RTD) between the first 50 and 100ms of isometric elbow flexion. Moreover, since the first 50ms of MVC are intimately related with the individual ability to achieve high levels of neural activation at the onset of muscle contraction (Andersen and Aagaard, 2006; Folland et al., 2014), we also provide preliminary evidence that both protocols induce a marked acute reduction in the neural component of the RTD. It is interesting to note that, even though LIBFR was performed at a lower volume load than HI exercise (~ 50%), the level of decrement in isometric MVC and the RTD was similar between protocols. This suggests that LIBFR is more efficient than HI exercise for eliciting fatigue. In line with this, past findings have also shown that, after individualizing BFR to a given percentage of AOP and setting exercise volume to a non-exhaustive regimen of 30+15+15+15 repetitions, LIBFR is as fatiguing as HI knee-extension as well as elbow-flexion exercise (Fatela et al., 2017; Kim et al., 2017). Thus, by showing that this effect is also extensive to the RTD obtained during maximal isometric elbow flexion, our findings extend those of previous studies.
Past data indicate that fatigue exerts a rapid and pronounced reduction in the early phase of explosive torque production and that this is secondary to the impaired neural drive (Buckthorpe et al., 2014). Considering this concept, central fatigue is the most likely cause of the RTD decrease during the first 50ms of MVC post-HI exercise and LIBFR. Under these circumstances, fatigue may be the reflection of an attenuated ability to produce doublet discharges, a reduced initial firing rate of motoneurons or even an increased recruitment threshold of motor units (Binder-Macleod and Barrish, 1992). There is unequivocal evidence that metabolic stress (i.e., depletion of phosphocreatine, increased accumulation of protons and lactate), which is highly heightened via LIBFR as well as through HI exercise, can elicit significant levels of fatigue and impose a high neuromuscular demand (Dankel et al., 2017a; Fatela et al., 2017; Husmann et al., 2018). Further supporting this notion, it has been shown that the restricted blood flow promotes the development of central fatigue after 4 sets of dynamic knee extensions at 30% 1RM (Husmann et al., 2018). Although several mechanisms may underlie the reduction in voluntary activation post-LIBFR and HI exercise, one mechanism that presumably accounts for decreased motoneuron firing rates under conditions of metabolic stress is the inhibitory feedback of group III/IV muscle afferents at the supraspinal level (Butler et al., 2003; Martin et al., 2008). Such a relationship is reinforced by the findings of Debold (2012) showing that acute reductions in RTD values are strongly related to an accumulation of H+.
Limitations
This study has several limitations. First, the RTD was only measured at 600 of elbow flexion. It has been shown that the isometric force-producing capabilities of muscles (RTD and peak torque) vary as a function of the joint angle (Murphy et al., 1995). Thus, the present findings cannot be extrapolated to joint angles other than 600 of elbow flexion. Nevertheless, since testing was consistently performed at the joint angle compatible with peak torque for the movement of interest (Murphy et al., 1995), it should be emphasized that we followed the recommendations of the available literature on measuring the RTD using isometric contractions (Yang et al., 2014). Second, instead of prescribing all sets to volitional failure, we pre-defined a standardized number of repetitions to all participants in both protocols. According to a recently published opinion on muscle hypertrophy (Dankel et al., 2017b), this approach may lead to unfair comparisons between protocols because the levels of fatigue at exercise termination may not be homogenous across conditions. Nevertheless, from a practical standpoint, performing blood-flow restricted exercise to volitional failure is not recommendable as it exacerbates delayed onset muscle soreness, which potentially limits training frequency (Fahs et al., 2012). To avoid these negative effects, LIBFR has been widely prescribed using the 30+15+15+15 repetitions protocol; an alternative that is highly effective in enhancing acute muscle activation and increasing muscular strength as well as hypertrophy without increasing indices of muscle damage (Loenneke et al., 2014; Scott et al., 2015). Thus, our methodologic approach to the prescription of LIBFR is well supported by past research. Third, previous research has shown that both the absolute and relative RTD are lower during isometric compared to dynamic test contractions (Pryor et al., 1994). Thus, based on our data, we cannot exclude possible differences between the effects of HI exercise and LIBFR on the dynamic RTD. Fourth, it is well known that the influence of neural and intrinsic contractile properties of the RTD changes throughout the force-time curve (Andersen and Aagaard, 2006; Folland et al., 2014). Connective tissue stiffness has also been shown to be predictive of the RTD during the first 100 ms of isometric torque production (Bojsen-Moller et al., 2005). We did not record electromyographic data, nor evoked twitch or octet contractions in this study. The mechanical properties of the biceps brachii tendonaponeurosis complex were also not measured. Thus, despite eliciting similar magnitude of effect on the sequential RTD, we cannot determine whether both protocols of resistance exercise affect all these determinants of explosive force similarly. Finally, we only measured the immediate impact of both protocols on MVC and the RTD. Past research indicates that recovery from HI and LI resistance exercise follows a different time course (Carroll et al., 2017). To our knowledge, the time course of recovery from neuromuscular fatigue has not yet been compared between HI exercise and LIBFR. However, differently from that seen in response to HI resistance exercise, there is preliminary evidence that the acute changes in torque following LIBFR recover within 1 h post-exercise (Carroll et al., 2017; Loenneke et al., 2013). Thus, our findings cannot be generalized to time points other than the immediate post-exercise recovery.
Conclusions
In conclusion, the present study showed that, when prescribed for a predefined number of repetitions, both LIBFR and HI exercise induced similar acute attenuation of the absolute RTD up to the first 100 ms of MVC. Importantly, these results were obtained in a context of different volume loads between protocols (the volume load of HI exercise almost doubled that of LIBFR). We also found that the reduction in the RTD between 50-100 ms (in response to both protocols) was largely explained by an acute deficit in muscle strength at the immediate post-exercise time point. In contrast, as supported by our data on the relative RTD, the negative impact of each protocol on the first 50 ms of rising muscle torque did not depend on lower levels of muscle strength after exercise. Thus, from a practical standpoint, we contend that LIBFR may be more efficient than HI exercise for affecting the neural component of the RTD.